
Annotated list of publications and presentations
Each of the entries below is followed by a brief personal comment.
Reading up from the bottom, this list charts the trajectory of my research career from graduate school through retirement.
Written with benefit of hindsight, the comments also map the origin and subsequent history of the "evolutionary tuning knobs" metaphor.
before 1981 | 1981-1990 | 1991-2000 | 2001-2010 | 2011-2020 | since 2020 | Epilogue
My career has been ultimately inspired by my interest in natural history. Since childhood, this interest has stimulated a desire to understand patterns of structure and genetics underlying adaptation, including the marvellous evolutionary malleability of animal behavior (cf. King 2013). I have grown ever more impressed by "extremely minute parts so shaped and situated as to form a marvelous organ" (to quote 17th-century microscopist Marcello Malpighi), especially by the exquisitely organized complexity of the nervous system (whose marvelous cells Santiago Ramón y Cajal memorably called "mysterious butterflies of the soul"). In each generation, such complexity somehow emerges anew from a single fertilized egg.
For me, the concept of individually identifiable nerve cells, which I first encountered during my initial weeks in graduate school, has been particularly influential.
Most of the peer-reviewed publications listed below are available somewhere on the internet, at least in abstract; full-text PDFs may be requested by email. Meeting presentations (posters and slide shows; not peer-reviewed) are by nature ephemeral (except for the pdfs archived here).

(2025b) King, D.G. Evolvability and lineage selection. Science E-Letter (published 19 March 2025).
This contribution is an e-letter (not appearing in print) commenting on a research article by Barnett, et al., Science 21 February 2025, Experimental evolution of evolvability. It appears farther down on the same webpage as Barnett et al.'s article. From my point of view, the principal virtue of this piece is that it appears in a fairly prominent venue and cites my two 2024 publications (below).(This short note might betoken the end of my publishing career, unless unexpected inspiration overtakes the growing limitations of advancing age.)

(2025a) King, D.G., Tandem repeat polymorphisms shape local adaptation. Trends in Ecology & Evolution 40: 331-334, https://doi.org/10.1016/j.tree.2025.02.004.
This essay highlights a report exploring "the extent to which STRs, especially those that that are linked to adaptive traits, could be shaped by the environment" [Ranathunge and Welch (2024) Clinal Variation in Short Tandem Repeats Linked to Gene Expression in Sunflower (Helianthus annuus L.), Biomolecules 14:944 https://doi.org/10.3390/biom14080944]. The authors have kindly shared with me that their report, the culmination of a multi-year project including several related articles, was inspired in part by two 1997 articles by Kashi, Soller, and myself.
My research career began with ecology, with participation in an NSF-sponsored program while I was still in high school. So I was delighted to finally land a paper in Trends in Ecology & Evolution. My proposal for this paper was my fourth attempt to publish in the journal; previous attempts were eventually accepted into Endeavour (1997) and Trends in Genetics (2006, 2024).
(2024b) King, D.G., Evolving a favorable distribution for mutation effects. Trends in Genetics 40:819-821. doi: 10.1016/j.tig.2024.07.009. [email for PDF: dgking@siu.edu].
This essay was was written with the intent of calling attention to my just-published article in The Journal of Physiology (below). It is a much shorter, more readily digestible version of that article, written for a venue where hopefully it is more likely to be noticed by geneticists. Perhaps it might also encourage someone to explore the potential of indirect selection through computational modelling of population genetics. I wish I could have offered a citation for Ranathunge and Welch's 2024 report in Biomolecules, but their paper appeared too late for inclusion.
(2024a) King, D.G., Mutation protocols share with sexual reproduction the physiological role of producing genetic variation within 'constraints that deconstrain'. The Journal of Physiology 602: 2615-2626; doi.org/10.1113/JP285478. [abstract] [free-access pdf]
Unexpectedly, after nine years of retirement, I stumbled into an opportunity to write for a special "physiology of evolution" issue of the venerable Journal of Physiology. The resulting essay offers a more substantial review of material explored on several previous occasions (listed below, especially posters in 2014 and 2015, also abstracts from 1985 and 2010), but this time written for J. Physiology readership.
Some readers might find it incredible that the outdated views of mutation addressed in this essay might still need refutation. Yet even in 2023, in Nature Reviews Genetics 24:3, we can read that "Sturtevant's [1937] views on the evolution of mutation rates (see 2014, below) are likely to continue to influence future efforts to understand what remains a fundamental question in biology." My primary impetus for this ms. was to continue challenging just such misleading but persistent influence.
This article is effectively an updated manifesto, for ideas that I first put into writing forty years earlier, in 1983.

(2015) King, D.G., Might constrained mutability be as advantageous as sex? (In this pdf, the poster display follows the "Further reading" list.)
This poster, like the two immediately below, takes direct aim at the classic but misleadingly simplistic assertion, famously declared by George Williams in 1966, that "[N]atural selection of mutation rates has only one possible direction, that of reducing the frequency of mutation to zero... So evolution takes place, not so much because of natural selection, but to a large degree in spite of it."
(For several years, I thought that this post-retirement meeting presentation would represent my final effort to advocate for the idea of mutation protocols as metaptation. This was somewhat disappointing, since posters are ephemeral (except as archived here). However, in 2023 I was invited to contribute an article (2024a) to a special issue on "physiology of evolution" in The Journal of Physiology. A couple closely related articles followed in shorter order [2024b, 2025a].)
(2014) King, D.G., A lesson from sex: Abundant variation can be worth its high price. (In this pdf, the poster follows an abstract and reading list.)
This poster (presented for the Society for Molecular Biology and Evolution annual meeting in San Juan, Puerto Rico), like the ones immediately above and below, challenges the misguided presumption that, in Sturtevant's oft-quoted words from 1937, "the vast majority of mutations are unfavorable... [M]utations are accidents, and accidents will happen." But this presumption ignores the simple fact that sexual reproduction is, fundamentally, a vast source of potentially advantageous mutations: Practically every instance of meiotic recombination creates DNA sequences that have never previously existed. And nearly all of these are NOT unfavorable.
☞ Simply because they are seldom deleterious, the products of reciprocal recombination are often explicitly excluded by definition from textbook descriptions of mutation. For example, in the glossary of one highly regarded textbook (Douglas Futuyma, Evolutionary Biology 3rd ed., 1998), "mutation" is actually defined as any alteration of the genome "that is not manifested as reciprocal recombination [emphasis added]."
(2013b) King, D.G., Whence variation? Rethinking mutation and evolvability. (In this pdf, an abstract and a reading list follow the poster.)
This poster, together with the two immediately above, takes direct aim at the classic but misguided dogma, reiterated as recently as 2012 by Elmore et al. (Genes Genomes Genetics 2: 1643-1649), that "a well established and supported tenet of evolutionary theory is that, because most new mutations are deleterious, selection in all organisms will act to reduce mutation rate toward the physiology- or selection-imposed minimum." A more thorough and much more expansive refutation followed in 2024
(2013a) King, D.G. What can giant axons tell us about genetics and evolution? International Society for Neuroethology Newsletter, July/August 2013, pp. 5-7.
A pair of giant axons in a small fly
(note additional large-axon pairs).As my retirement approached, I prepared this essay -- illustrating distinctive patterns of nerve fibers in several species of flies -- as a "swan song" to my interest in specialized neuronal form and function. This essay encapsulates the logic which has guided my career for several decades.
☞ The idea of individually specialized nerve cells has fascinated me since 1970, when I was introduced to it during my first "rotation" in graduate school in Ted Bullock's lab at UCSD. There I was assigned to record electrical activity from one particular cell (R-15, "the parabolic burster") in the abdominal ganglion of Aplysia.
My doctoral dissertation (1976b and 1976c, below) focused on several individually-identifiable cells in the stomatogastric ganglion of the California spiny lobster. My postdoctoral work was directed toward understanding unique giant axons in Drosophila. During the early 1980s, I began examining a variety of different fly species in hopes of pinning down the phylogenetic origin of such curious giant axons. (Although a simple story of giant fiber evolution continues to elude me, the specimens collected for that purpose served admirably for my 1991 report on the evolutionary origin of the dipteran cardia.)
Hoping to stimulate more widespread interest in the evolution of individually-identifiable nerve cells, I reported further details of fly neurons at several annual meetings of the Society for Neuroscience. When those presentations received very little attention, I put neurobiology aside in favor of my transition to studies of evolutionary genetics. Yet even this transition was indirectly driven by my desire to understand how individually-identifiable nerve cells could evolve, particularly when the totality of information in a genome would seem inadequate (at least to me) to specify such precision circuitry.
This essay expresses my conviction that the goal of understanding nervous system organization might benefit by addressing evolutionary processes. (For a related, extended discussion, see my 2001 online-only essay, "Evolution of neural circuits." Also cf., 1988b, 2007c.)
| ||||
(2012) King, D.G. Indirect Selection of Implicit Mutation Protocols. Annals of the New York Academy of Science 1267:45-52. doi: 10.1111/j.1749-6632.2012.06615.x [author preprint] [email for PDF: dgking@siu.edu]
At the time, this essay felt like the culmination of my work on the implications of indirect selection for evolutionary processes. Subsequent poster presentations, at meetings held in 2013, 2014, 2015, and also listed above, have offered some further elaboration. A dozen years later I found opportunities for more forceful presentations, published in 2024.
(2011d) King, D.G. Indirect selection of local mutation rates (abstract only). DIMACS Conference on Effects of Genome Structure and Sequence on the Generation of Variation and Evolution, August 9 - 11, 2011.
In 2011 I was invited to participate in a conference at the Rutgers University Center for Discrete Mathematics and Theoretical Computer Science. My abstract, initially published in proceedings of this conference, was subsequently expanded into an essay (2012, above), published in a collection edited by Lynn H. Caporale with the same title as the conference, "Effects of Genome Structure and Sequence on the Generation of Variation and Evolution," in the Annals of the New York Academy of Sciences, Volume 1267 (2012).

(2011c) King, D.G. Genetic Variation Among Developing Brain Cells. Science E-Letter (published 16 May 2011).
This letter-to-the-editor (continuing our campaign to publicize the evolutionary role of repetitive DNA) was submitted in response to a news article by Gretchen Vogel ( Science 332:300-301, 2011), "Do Jumping Genes Spawn Diversity?" (This is an e-letter, not appearing in print; it appears farther down on the same webpage as Vogel's article.)
(2011b) King, D.G. Light on genetic dark matter (letter to the editor). Science News 179(7):30 (26 March 2011).
Another letter-to-the-editor in our campaign to publicize the evolutionary role of repetitive DNA, this time in a science magazine published for the educated general public.
(2011a) King, D.G. Evolution of simple sequence repeats as mutable sites. In: Anthony Hannan, ed., Tandem Repeat Polymorphisms: Genetic Plasticity, Neural Diversity and Disease, Landes Bioscience and Springer Science+Business Media. Adv Exp Med Biol. (2012) 769:10-25. [preprint] [email for PDF: dgking@siu.edu]
I was invited to submit this essay by the editor, A. Hannan, after previously inviting him to share the authorship of a 2008 Trends in Neuroscience review. Unconstrained by any publisher-imposed word limit, in this article I was able to develop a more thorough consideration of simple sequence repeat evolution than I had yet accomplished anywhere else (cf. 2012).
Here, in addition to reviewing both classical and more-nuanced discussions of natural selection for mutation-rate, I consider the significance of the phylogenetic distribution of tandem-repeat sites, many of which are conserved across vast spans of evolutionary time. Such conservation comprises evidence, which I find especially compelling, that the evolution of tandem repeats has indeed been driven by more than mutation pressure and genetic drift. Positive (albeit indirect) selection -- favoring, preserving, and shaping the mutagenic potential of tandem repeats -- has surely been involved as well.
(Due to citations added in proof, the author preprint available here differs slightly from the print version of this article.)
 |
| Image by Milo Winter,
Aesop's Fables, 1919 (adapted from Wikipedia Commons) |
(2010b) King, D.G. Metaptation: Metaphors for genome evolution. In: Structural and Functional Diversity of the Eukaryotic Genome, Verhandlungen des naturforschenden Vereines in Brünn, Specialausgabe, p. 40.
This abstract, which offers an updated definition for the word "metaptation" (first introduced in 1985), was written for an international workshop held at the Mendel Museum in the former St. Thomas Abbey (Brno, Czech Republic) where Gregor Mendel grew his peas. It might be my only publication to include an exclamation point!
My invitation to speak at this workshop came from Ed Trifonov, who had been coauthor on our 2006a article in The Implicit Genome. The image at right links to the slideshow for my presentation. Most of the other presentations at this meeting were dense with molecular information that went way over my head. Cheeky of me, a neuroscientist, to speak in Mendel's abbey at a molecular genetics seminar.
Quite a few years later, I took advantage of my post-retirement 2024 Journal of Physiology paper to slip the word "metaptation" into mainstream biology literature.

(2010a) King, D.G. A hypertext resource for self-directed learning. (poster pdf)
Throughout my career, my principal teaching assignment has been histology. In 2000 I began building a website emulating my role as instructor, to help students learn that subject. This poster describes the resulting resource.
The website itself, consisting of several hundred webpages of extensively hyperlinked text and images, was initially posted at a time when internet resources were not as extensive as they have since become. (This website remains a work in progress.) By 2010 this website had acquired sufficient popularity that a Google search for the single word "histology" returned this site in first place out of several million hits. For obscure reasons it was deleted from the SIU School of Medicine webserver in 2021; nevertheless, after being resurrected at its current URL, histology.siu.edu, this website has regained its visibility to Google searching. I have been extremely gratified to receive "fan mail" from around the world, expressing appreciation for the quality of this site.
[Incidently, the gold medallion at right was a science-fair prize I received in junior high school, long before histology became central to my professional career. That this medallion illustrates a microscope is purely coincidental. Or maybe it was a message from the future!]In 2021, I began working on a new page for this website, one dedicated to the history of histology as told through histology eponyms. This page eventually grew to be quite lengthy, with over 80 entries. I very much enjoyed tracking down material illustrating the lives of so many pioneers who helped develop the science of tissue structure, specially since this is the science that eventually became the foundation for my own career.
(2009b) King, D.G., Why do so many neurodevelopmental genes contain repeat-based mutable sites? (poster pdf)
I prepared this poster for the 2009 meeting of the Society for Neuroscience (the last such meeting which I have attended), hoping to call some attention to especially intriguing observations of tandem-repeat conservation in two papers published that same year.

(2009a) King, D.G., and Y. Kashi Heretical DNA sequences? Science 326: 229-230.
This letter-to-the-editor was written to challenge simplistic application of the "conservation equals function" paradigm in evolutionary genetics, while publicizing the intriguing results recently published by Riley & Krieger (cf., 2009 poster).
(2008b) King, D.G., Facilitating Evolution: Adaptation and Simple Sequence Repeat Polymorphism. (poster pdf)
This poster was prepared for an ecology symposium; an abstract and a reading list follow the poster.
 (2008a)
Fondon III, J.W., E.A.D. Hammock, A.J. Hannan, and D.G. King Simple sequence repeats: Genetic modulators
of brain function and behavior. Trends
in Neurosciences 31: 328-334. doi: 10.1016/j.tins.2008.03.006.
[authors' prepublication MSWord
document] [email for PDF: dgking@siu.edu]
(2008a)
Fondon III, J.W., E.A.D. Hammock, A.J. Hannan, and D.G. King Simple sequence repeats: Genetic modulators
of brain function and behavior. Trends
in Neurosciences 31: 328-334. doi: 10.1016/j.tins.2008.03.006.
[authors' prepublication MSWord
document] [email for PDF: dgking@siu.edu]
This review emphasizes the applicability of our tuning-knob hypothesis (1997b, 1997c, 2006a) in the domain of neuroscience. My principal roles in the preparation of this article were recruiting coauthors with expertise on the effects of simple sequence repeat variation, editing the tens of thousands of words provided by these coauthors into the word-limit imposed by the journal, and writing the introduction and conclusion.
(2007c) King, D.G., What's so special about giant fibers? (slide show pdf)
This slide show was prepared for a symposium on myelin (cf. evolution of myelin), at the National Evolutionary Synthesis Center (the precursor to TriCEM). Content is similar to 2013a (above). My invitation to this symposium came from Dan Hartline, who had been a professor down the hall from Allen Selverston's laboratory at UC San Diego while I was a graduate student in that lab.
(2007b) King, D.G., and Y. Kashi Mutation rate variation in eukaryotes: evolutionary implications of site-specific mechanisms. Nature Reviews Genetics 8 (November 2007) | doi:10.1038/nrg2158-c1.
This is the second of two short essays published in 2007, in response to published papers which failed to address overwhelming evidence for extensive variation in localized, site-specific mutation rates. Unfortunately, the significance of such sites was still being ignored years later (e.g., Zhang 2022, also in Nature Reviews Genetics). Mutation rate continues to be treated as a single, unitary parameter, while Sturtevant's obsolete 1937 dictum -- that "mutations are accidents, and accidents will happen" -- persists as misleading dogma (cf., 2012, "Indirect selection...," also 2024, "Mutation protocols...").
(2007a) King, D.G., and Y. Kashi Mutability and Evolvability: Indirect selection for mutability. Heredity 99:123-124. doi:10.1038/sj.hdy.6800998. [email for PDF: dgking@siu.edu]
This is the first of two short essays published in 2007, in response to published papers which failed to address overwhelming evidence for extensive variation in localized, site-specific mutation rates. Unfortunately, the significance of such sites is still being ignored years later (e.g., Zhang 2022). Mutation rate continues to be treated as a single, unitary parameter, while Sturtevant's obsolete 1937 dictum -- that "mutations are accidents, and accidents will happen" -- persists as misleading dogma (cf., 2012, "Indirect selection...," also 2024, "Mutation protocols...").
(2006e) King, D.G., Facilitating Evolution: Simple Sequence Repeats Make Genes Adjustable. (poster pdf).
While attending a conference at UC Berkeley to present this poster, I shared a hotel room with John Fondon who had made a splash in 2004 by reporting the role of repetitive DNA in morphological variation of dogs. I later recruited Fondon as a coauthor for our Trends in Neuroscience paper (2008).
(2006d) Y. Kashi and D.G. King Has simple sequence repeat mutability been selected to facilitate evolution? Israel Journal of Ecology and Evolution 52:331-342. [Abstract] [email for PDF: dgking@siu.edu]
This essay continued our campaign to publicize the evolutionary implications of simple sequence repeats.
(2006c) Hammock, E.A.D., and D.G. King Genes and neuroethology: How can evolution adjust behavior? International Society for Neuroethology Newsletter, November 2006, pp. 4-7.
This newsletter article highlighted new information relating simple sequence repeats to affiliative behavior in voles, as reported by Elizabeth Hammock in the previous year. I later recruited Hammock as a coauthor for our Trends in Neuroscience paper (2008a).
(2006b) Y. Kashi and D.G. King Simple Sequence Repeats as Advantageous Mutators in Evolution. Trends in Genetics 22: 253-259. [Abstract] doi: 10.1016/j.tig.2006.03.005 [email for PDF: dgking@siu.edu]
This review updated our 1997 TiG review, with greater emphasis on evolutionary implications. It has become our most highly-cited publication.
☞ Evidence for such "advantageous mutators" has continued to accumulate. Some of the more recent publications are cited in my 2024 articles in The Journal of Physiology and Trends in Genetics, which generalize the concept to additional protocols for mutation besides tandem repeats.

(2006a) King, D.G., E.N. Trifonov, and Y. Kashi Tuning Knobs in the Genome: Evolution of Simple Sequence Repeats by Indirect Selection. In: Lynn H. Caporale, ed., The Implicit Genome, Oxford University Press. [Abstract] [Table of contents for The Implicit Genome] [email for preprint: dgking@siu.edu]
Lynn Caporale offered the invitation to submit this essay, based on mutual interests established at her 1998 NYAS meeting, "Molecular strategies of biological evolution" (1999b). In previous reviews coauthored with Kashi and Soller (listed below), I had depended almost entirely on those two coauthors' knowledge of the genetics literature. Preparing to write this review finally gave me enough confidence to continue writing on my own.
Another essay in this volume, "Epilogue: An Engineering Perspective: The Implicit Protocols," by John Doyle, Marie Csete, and Lynn Caporale, introduced the "protocols" metaphor which I subsequently adapted into my 2012 essay in Annals NYAS.
(2002) King, D.G., and J.G. Killian Binocular rivalry, bipolar disorder, and aging: Perceptual alternation rate correlates with age as well as with psychiatric diagnosis. Program No. 803.4 Orlando FL: Society for Neuroscience. [Abstract with graph of data] [extended discussion]
Prompted by events in my personal life, I put aside my principal interest in evolutionary genetics for a three-year project summarized in this abstract. The project was funded by the Brain & Behavior Research Foundation (which I had learned about through membership in NAMI). Its subject, binocular rivalry, is a curious phenomenon with implications for the neural substrate of consciousness (more). Killian, a graduate student at the time, gave up a secure job to assist with this project.
(2001) King, D.G. Evolution of neural circuits. SIUC Cognitive Science Colloquium.
This text of a seminar presentation on October 5, 2001, summarizes most of the central ideas of my research career, both before and after this date.
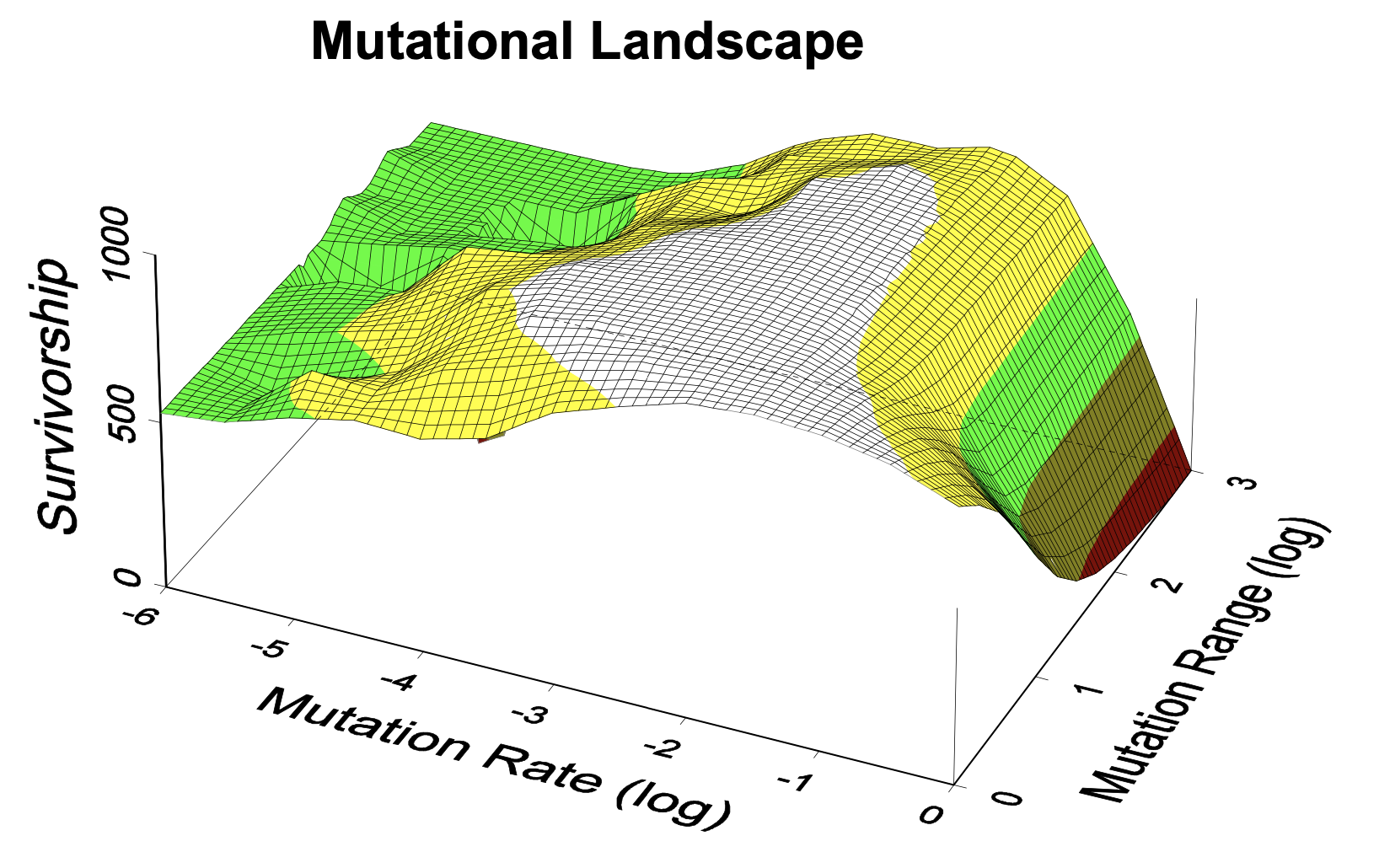
(2000) King, D.G. Indirect selection on the mutational landscape: An evolved role for the mutability of repetitive DNA? Poster #218, in Evolution 2000, (ed. M.J. Wade), Proceedings of the 2000 annual meeting of the Society for the Study of Evolution. [extended abstract, pdf]
An extended abstract of this meeting presentation, including graphically illustrated results of a simple model of population genetics, was published in a CD-ROM of the proceedings of the 2000 meeting of the Society for the Study of Evolution. (Unfortunately, the full contents of this CD do not appear to be available online.) Disappointingly, this poster presentation garnered essentially no interest; reaching a broad audience with this idea has proven challenging (cf. 2007a and 2007b, above).
(1999c) Krajewski, C., Fain, M., L. Buckley and King, D. G. (1999) Dynamically heterogenous partitions and model-based phylogenetic inference: An evaluation of analytical strategies with cytochrome b and ND6 gene sequences in cranes. Molecular Phylogenetics and Evolution 13: 302-313. [Abstract]
For this and a 1996 paper also coauthored with Krajewski, my only contribution was the coding of conceptually-simple computer programs which facilitated the comparison of DNA sequences. Matt Fain and Larry Buckley were Krajewski's graduate students.
(1999b) King, D.G. Modelling selection for adjustable genes based on simple sequence repeats. In: Molecular Strategies in Biological Evolution, Annals of the New York Academy of Science 870: 396-399.
This abstract was published in the proceedings of a 1998 meeting of the New York Academy of Sciences, "Molecular strategies in biological evolution," organized by Lynn Helena Caporale. Prior to this meeting, I had written a computer program simulating a very simple model of population genetics. Output from many runs of that program had convinced me that the speculative idea of "indirect selection" could plausibly occur as a real phenomenon. (A more elaborate version of this computer model was prepared for the 2000 meeting of the Society for the Study of Evolution.)I travelled to this meeting not knowing what to expect. At the time I still felt like a naive outsider in the field of molecular evolution. But at this meeting I found a small community of biologists who shared my growing interest in molecular mechanisms that could plausibly function as strategies for evolution. My participation in this meeting was a turning point in my career, as I gained confidence to continue pursuing the "evolutionary tuning knobs" concept. (Incidently, my flight from Memphis to New York to attend this meeting had memorably passed through a spectacular "cathedral" of stormclouds, as our airliner wove closely among columns of thunderheads that stretched for miles into the distance. In hindsight this was a remarkable omen for subsequent events in my career.)
I was startled (very pleasantly so!) when the first speaker (James A. Shapiro) cited our recently published Trends in Genetics review (1997b). I was astonished when a subsequent speaker, Ed Trifonov, suggested that a plausible metaphor for the behavior of simple sequence repeats might be that of "tuning knobs." When I approached him after his talk with a reprint of our "Evolutionary Tuning Knobs" essay (1997c), his surprised reaction was priceless. We later collaborated on a 2006 essay for Caporale's volume, The Implicit Genome.
Delightfully (for me), my own participation at this meeting was reported in a "News Focus" article for the journal Science: "How the Genome Readies Itself for Evolution" (E. Pennisi, Science, 21 Aug 1998, Vol 281: 1131-1134, DOI: 10.1126/science.281.5380.1131). By summarizing several of the presentations at this meeting, Pennisi's article provided a nice overview of a newly emerging field of genetics, one focussed on studying the molecular basis for evolvability. While attending this meeting, I hadn't yet appreciated how extensively I myself would continuing publishing in this field.
 (1999a) King, D.G., and M. Soller Variation and fidelity: The
evolution of simple sequence repeats as functional elements in adjustable genes. In: S.P. Wasser, ed.,
Evolutionary Theory and Processes: Modern Perspectives, pp. 65-82. Kluwer
Academic Publishers, Dordrecht, The Netherlands. [Abstract]
[email for PDF: dgking@siu.edu]
(1999a) King, D.G., and M. Soller Variation and fidelity: The
evolution of simple sequence repeats as functional elements in adjustable genes. In: S.P. Wasser, ed.,
Evolutionary Theory and Processes: Modern Perspectives, pp. 65-82. Kluwer
Academic Publishers, Dordrecht, The Netherlands. [Abstract]
[email for PDF: dgking@siu.edu]
This essay, published in a festschrift dedicated to Eviatar Nevo, continued our nascent campaign to publicize the evolutionary implications of simple sequence repeats.
(1998) King, D.G. Cells "R" Us. Brain Awareness Week at SIUC.
This entry is the text of a public lecture offered during a program of public educational activities sponsored by the Society for Neuroscience, and the Dana Foundation. My goal in this presentation was to acquaint attendees with the basic idea that nerve cells, as the working parts of the brain, are intimate participants in sensation and movement.
(1997d) King, D.G. Is there a role for comparative genetics in neuroethology? International Society for Neuroethology Newsletter, July 1997.
This newsletter article was effectively a preview of subsequent 2006c and 2013a articles in the same society newsletter.

(1997c) King, D.G., M. Soller, and Y. Kashi Evolutionary tuning knobs. Endeavour 21: 36-40. [publisher pdf] [email for PDF: dgking@siu.edu]
This essay properly introduced "tuning knobs" as a metaphor for the functionality of repetitive DNA (first mentioned in abstract two years earlier, in 1995). This same metaphor had been independently suggested by Ed Trifonov, at a 1998 meeting of the New York Academy of Sciences ("Molecular strategies in biological evolution," organized by Lynn Helena Caporale). Trifonov's surprise when I showed him a reprint of this article led eventually to our collaboration on an essay published in 2006a, "Tuning knobs in the genome: Evolution of Simple Sequence Repeats by Indirect Selection." A few years later he invited me to participate in a conference at Gregor Mendel's abbey in Brno.I was quite pleased when the "evolutionary tuning knob" concept was picked up in an article in Scientific American (January, 1999, pp. 94-99, "DNA Microsatellites: Agents of Evolution?)" by Richard Moxon and Chris Wills, who also suggested the equivalent metaphor of "rheostat."
☞ I had initially encountered the metaphor of "twiddling a knob on a concept" in an essay by Douglas Hofstadter, "Variations on a theme as the crux of creativity" when his essay first appeared in Scientific American in October, 1982. (This essay was subsequently reprinted in Metamagical Themas: Questing for the Essence of Mind and Pattern, Basic Books 1985.) Variations on a theme are commonplace in biology. At that time, while I was developing the idea of metaptation, I had begun to wonder how a genome might quantitatively explore variations on a theme. I learned of a plausible answer in a 1983 essay, "Updating the theory of mutation," by Drake, Glickman, & Ripley (American Scientist 71:621-630) which described the peculiar mutational characteristics of repetitive DNA. But it took another decade for evidence to appear that DNA could indeed have such a function, leading first to my 1994 letter to Science and eventually to this "Evolutionary Tuning Knobs" essay.The concept that variation in simple sequence repeat number could "adjust" gene function emerged repeatedly in the early 1990s, and continued to reemerge thereafter. While preparing a manuscript in the early 2020s, I came across a paper that I had not previously noticed ("Engineering in Genomics: variable number tandem repeats as agents of functional regulation in the genome," G. Breen et al. [2008], Engineering in Medicine and Biology Magazine 27: 103-104, 108), presumably because it had not cited any of the reviews that had by then become familiar to me. This reminded me that different communities of biologists can sometimes remain fairly oblivious to one another. Indeed even the "tuning knob" / "rheostat" metaphor continues to reemerge, appearing anew in 2023, apparently independently from this history, in a featured article in Science (Horton, C.A. et al. [2023] "Short tandem repeats bind transcription factors to tune eukaryotic gene expression," Science 381, eadd1250).The "tuning knob" metaphor also features prominently in two essays published in 2024, in The Journal of Physiology and in Trends in Genetics, and another in 2025 in Trends in Ecology and Evolution.
 |
| Chezi and me (with Gregor Mendel) |
(1997b) Y. Kashi, D.G. King, and M. Soller Simple sequence repeats as a source of quantitative genetic variation. Trends in Genetics 13: 74-78. [Abstract] [email for PDF: dgking@siu.edu]
This review, my first collaborative publication with Morris Soller (Moshe) and his former student Yechezkel Kashi (Chezi), has become one of our most-cited publications (exceeded only by its sequel, 2006b). It helped establish the idea that simple sequence repeats could be significant sources of genetic variation.This collaboration had been prompted by my 1994 letter to Science. After noticing that letter, Moshe reached out to me from the Hebrew University in Jerusalem, proposing that we work together to publicize our shared idea. (Soller, coincidently, was an old friend of Ed Simon, my undergraduate genetics professor at Purdue; see my epilog.) To facilitate our long-distance correspondence, Moshe initiated me into the use of email. I finally visited Moshe in person in 2007, at his family home in Chicago. I met Chezi three years later, at a conference (2010b) held at Gregor Mendel's abby in Brno, Czech Republic.
(1997a) King, D.G. Behavioral adjustments [letter to the editor], American Scientist 85: 300-301.
This letter addressed an essay, "The evolution of arthropod nervous systems," which had been published in the journal's previous issue (Osirio, Bacon & Whitington, Amer. Sci., 85:244-253, May/June 1997). My letter posed a simple question: How can a limited genome support extraordinarily facile evolution of instinctive behavior? I then proposed that the answer might involve repetitive DNA. Alongside this letter, the journal published a response by the authors of the original essay, expressing strongly-worded skepticism. Such skepticism was common at the time (and still is, unfortunately), but was addressed that same year by our articles in Endeavour and Trends in Genetics (1997b, 1997c, above) as well as in several of our subsequent publications (e.g., 2008a), culminating in my 2013a essay for the International Society for Neuroethology. (For a related, extended discussion, see my 2001 online-only essay, "Evolution of neural circuits.")
(1996) Krajewski, C. and D.G. King, Molecular divergence and phylogeny: Rates and patterns of cytochrome b evolution in cranes. Molecular Biology and Evolution 13: 21-30. [Abstract]
Carey Krajewski is a friend and colleague at SIUC Zoology. For this and a subsequent 1999c paper also coauthored with Krajewski, my only contribution was the coding of conceptually simple-minded computer programs which facilitated the comparison of DNA sequences. (As simple as these programs were, I have heard that they've been used by other researchers, from The Smithsonian Institution in Washington DC to LaTrobe University in Australia.)
(1995) King, D.G., Triplet-repeat DNA: Unstable regulatory genes could be evolutionary "tuning knobs." (abstract only)
This abstract from a Society for Neuroscience annual meeting represents my first published mention of the "tuning knobs" metaphor, which was published more substantially two years later (1997c).

(1994) King, D.G. Triple repeat DNA as a highly mutable regulatory mechanism. Science 263: 595-596. [publisher pdf] [email for PDF: dgking@siu.edu]
This letter-to-the-editor (appearing in print just in time for my 46th birthday!) was prompted by a news article in Science ("The Puzzle of the Triple Repeats," 4 June 1993), which reported a peculiar quantitative relationship between severity of symptoms in Huntington's disease and number of CAG repeats in the causative gene. This relationship corresponded remarkably well with a speculation I had conceived a decade earlier, while developing the concept of "metaptation." By staking my claim to the hypothesis that repetitive DNA might confer upon genes the property of adjustability, this letter officially marked the beginning of my re-invention as a theoretical evolutionary geneticist. (Incidently, the opportunity to write this letter came during a dry spell for my research, when I was really unsure what to do next.)
I was over the moon when, just one week after Science published my letter, Science then published a report by Gerber et al. ("Transcriptional Activation Modulated by Homopolymeric Glutamine and Proline Stretches") which powerfully confirmed the plausibility of my hypothesis. I almost overlooked this confirmatory report, since its title consisted almost entirely of phrases that had not yet become part of my familiar working vocabulary.
Publication of this letter led directly to collaboration with Yechezkel Kashi and Morris Soller, of the Hebrew University in Jerusalem, who had independently developed similar ideas. Together we have coauthored several papers listed above, beginning in 1997 with Trends in Genetics and Endeavour (1997b and 1997c).
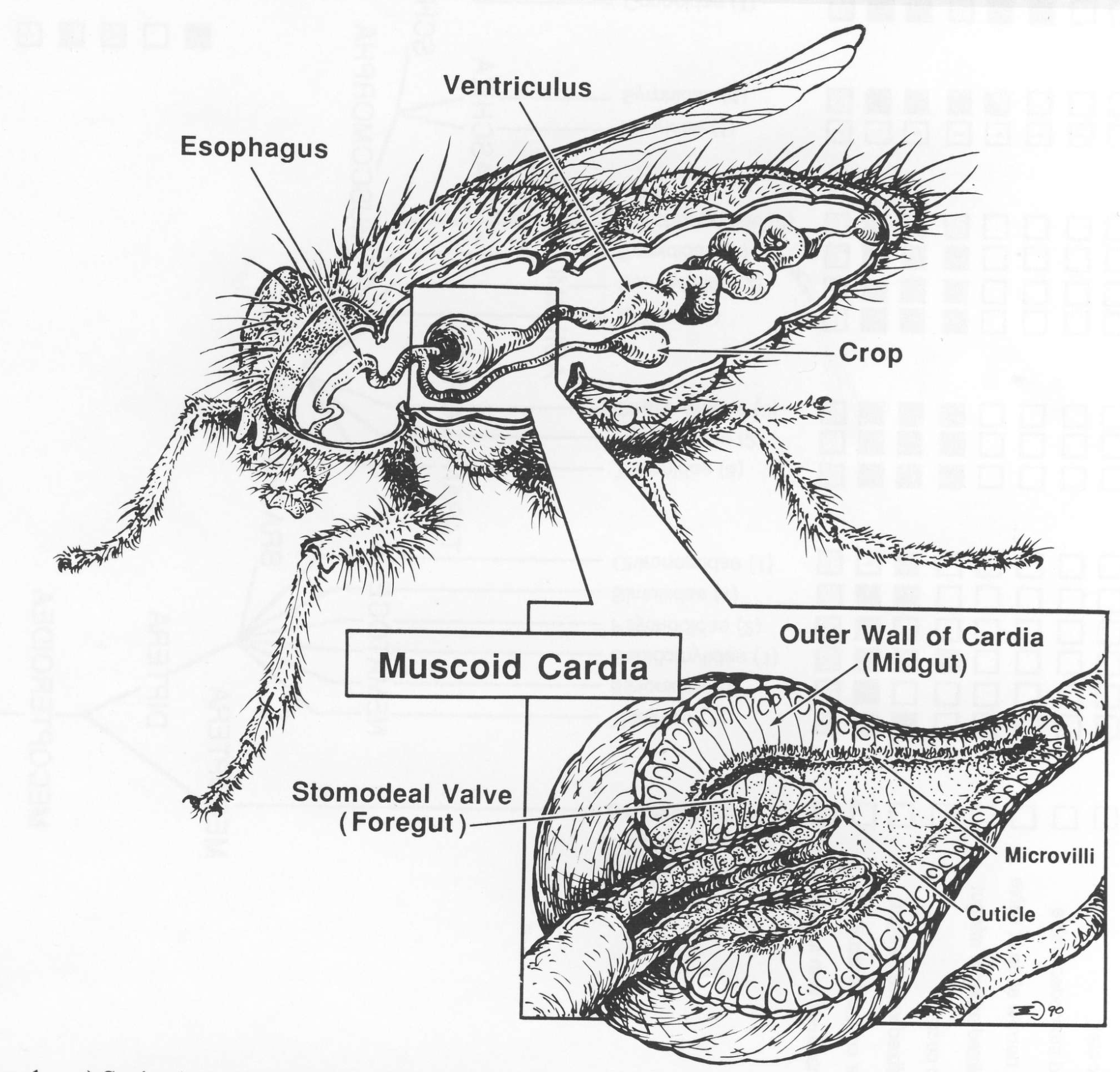
(1991) King, D.G. The origin of an organ: Phylogenetic analysis of evolutionary innovation in the digestive tract of flies (Insecta: Diptera). Evolution 45:568-588. [email for PDF: dgking@siu.edu]
Research leading up to this report was probably the most fun of any during my career. The project involved not only delightful fieldwork wandering around southern Illinois gathering specimens representing 65 distinct species, it also provided intense engagement with evolutionary thinking and phylogenetic reasoning. The outcome was a surprisingly elegant story of how a fairly complex organ had emerged. As a result, I became inordinately pleased with myself -- a neuroscientist by training -- for successfully publishing in a journal devoted to evolutionary biology. (I had not yet begun my full transformation from neuroscience into evolutionary genetics.) Although this report has not been cited nearly as often as several subsequent papers on molecular evolution (e.g., 1997b, 1997c, 2006b), it remains one of my favorite publications.
(1989) King, D.G. Phylogenetic diversity of cellular organization in the cardia of muscoid flies (Diptera: Schizophora). Journal of Morphology 202: 435-455. [email for PDF: dgking@siu.edu]
This publication set the stage for my 1991 report on the evolutionary origin of the dipteran cardia. It was based on a collection of various fly species initially intended for exploration of the phylogenetic origin of giant axons found in Drosophila, a study eventually published in the ISN newsletter in 2013a.
(1988b) King, D.G. On the evolution of specialized neuronal form: Phylogenetic variation in cervical giant axons of flies (Diptera). Proc. XVIII Int. Congress of Entomology, p. 70 (abstract only).
Travel to this Congress of Entomology meeting was especially memorable: My wife and I drove from southern Illinois to the meeting site in Vancouver, B.C., touring the country for five weeks with our son and daughter, ages 6 and 9.
Several years earlier (in 1980), I had described a giant-axon pathway in Drosophila. Shortly afterwards I had established that a homologous giant-axon pathway, although missing in tsetse flies (1983a), could nevertheless be found in other muscoid flies (1983b). In an effort to establish when this giant-axon pathway had first evolved, I set out to map its phylogenetic distribution by examining as many different flies as I could easily acquire. This provided me with a welcome excuse for outdoor fieldwork, collecting specimens around the delightful environs of southern Illinois. Eventually, by the time of this meeting, I had recorded axon size distributions in sixty-five species representing thirty-six families.Although observations presented at this Vancouver meeting did not suggest a simple story about the evolutionary origin of Drosophila giant fibers, they were nonetheless quite intriguing: Naive initial hypotheses were rejected one after another by new specimens. Axon size distributions among various fly species turned out to be unexpectedly diverse, even within a single family. A few images of this diversity were eventually published 25 years later. And the same collection of specimens served admirably for my analysis of the evolutionary origin of the dipteran cardia (in 1989 and 1991).
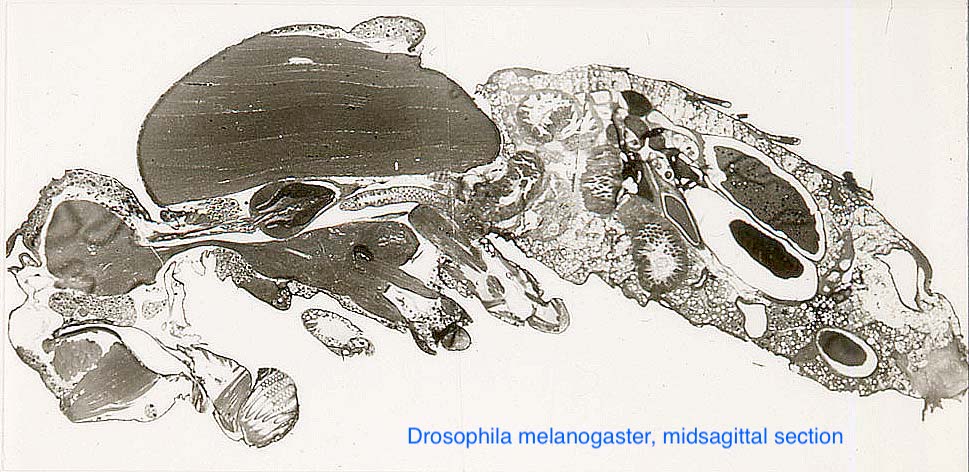
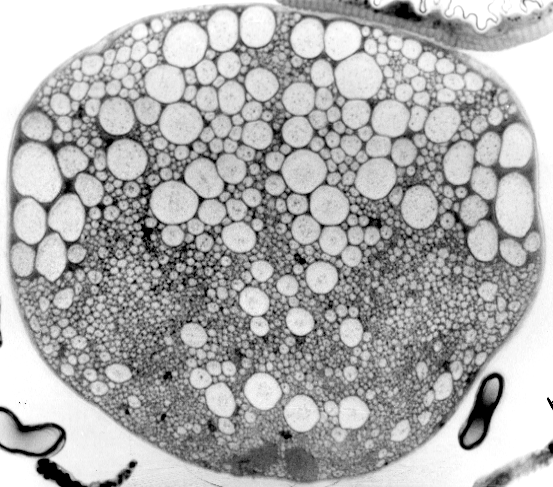
(1988a) King, D.G. Cellular organization and peritrophic membrane formation in the cardia (proventriculus) of Drosophila melanogaster. Journal of Morphology 196: 253-282. [Abstract] [email for PDF: dgking@siu.edu]
This study was prompted by accidental observation of abnormal cardia development in some behavioral mutants of Drosophila, whereupon I realized that this organ had never received a detailed histological description. This was my first substantial work that had not been grounded on work in the laboratories of my doctoral and post-doctoral advisors. It led directly to my 1989 and 1991 publications (above) on comparative anatomy of the Dipteran cardia.
I might never have paid attention to the fly cardia, but for its location atop the thoracic nervous system where I could not help but notice it while studying neural processes in the thoracic ganglion.
In a belated sequel, a study building on this description was published in 2011. The authors of that study had severely overinterpreted their results to draw conclusions which were grotesquely wrong in ways that would not be apparent without detailed understanding of the Drosophila cardia. Unfortunately, especially since those same authors had acknowledged my assistance identifying cell types in their micrographs, I became not only embarrassed that my own contribution might be clouded by association but more importantly distressed that the potential utility of the fly digestive system for research into gene expression might be permanently derailed by the appearance of such false information in the literature. Thankfully, in 2026 (38 years after my original description and 15 years since the erroneous conclusion had been published), a new report by Ewen-Campen et al. appeared which exposed the prior error and appropriately cited my work. The time course here was prolonged, but it was reassuring to observe the error-correcting process of science at work.
Especially with this 2026 report, I still hope my initial cardia publication, together with its two sequels (above), might serve to introduce the Dipteran cardia as a model system for studying evolutionary differentiation of cell function within a well-defined location.
(1985) King, D.G. Metaptation: A descriptive category for evolutionarily versatile patterns of genetic and ontogenetic organization. Abstract only, in Evolutionary Theory 7:222. [Abstract] [unpublished ms.]
This abstract marked the beginning of a major shift in my career orientation, from neuroscience to evolutionary genetics, culminating in a pre-retirement 2012 Annals NYAS paper and several post-retirement essays in 2024 and 2025, all on mutation protocols as exemplified by short tandem repeats.
The central idea lying behind this abstract -- that effective evolution might require "adaptations for adaptability" -- emerged while I was trying to understand what manner of genetic organization could possibly sculpt fine details of individually specialized nerve cells, such as Drosophila's giant axons.
"Metaptation," the title neologism here, is modelled after Gould & Vrba's coinage of "exaptation." This word has not yet entered mainstream usage, although it has been adopted by at least one researcher working with the evolvability of computer programs (Moshe Looks 2007, "Program evolution for general intelligence," full text). The idea is elaborated in William Calvin's book The River that Flows Uphill (1986, 2010).
Twenty-five years later, at a 2010b meeting in the Czech Republic, I presented a "new, improved" definition for the word. And I took advantage of my post-retirement 2024 Journal of Physiology article to slip the word "metaptation" into mainstream biology literature.
|
This 1985 abstract was originally written to accompany a lengthy essay that had been written in 1983 and submitted to a journal where it languished for two years before being rejected. (Meanwhile I had moved on with histological study of fly anatomy.) The original manuscript included my first recognition of the plausibility of "evolutionary tuning knobs," an idea that had been inspired by a 1982 essay by Douglas Hofstadter but not published properly until 1997.
☞ Here I must acknowledge a special debt to Steven Jay Gould, whose monthly essays in Natural History magazine inspired me to wonder about how evolution really works. Without the encouragement that Gould offered in his generous reply to a "fan letter," in which I shared some initial thoughts about evolvability, I probably would have lacked confidence to continue writing in a domain of biology where I had no formal training.
(1984) Wyman, R.J., J.B. Thomas, L. Salkoff, and D.G. King The Drosophila giant fiber system. In: Neural Mechanisms of Startle Behavior (R. Eaton, Ed.), Plenum Press, New York.
My inclusion as an author of this review, based on my participation as a postdoc in Robert Wyman's lab at Yale (1976-1977, with publication in (1980), was essentially honorary; I played no part in preparation of this book chapter for publication.
(1983e) King, D.G. Review of Genetic Neurobiology, by Hall, Greenspan, & Harris; in Quarterly Review of Biology, 58:286.
When I wrote this very brief book review (less than 1/2 page), genetics was still a fairly new tool for studying nervous systems. I have since come to believe that additional value can be obtained by using some of the more remarkable features of nervous systems as inspiration for re-thinking certain aspects of genetics (cf. What can giant axons tell us about genetics?, above).
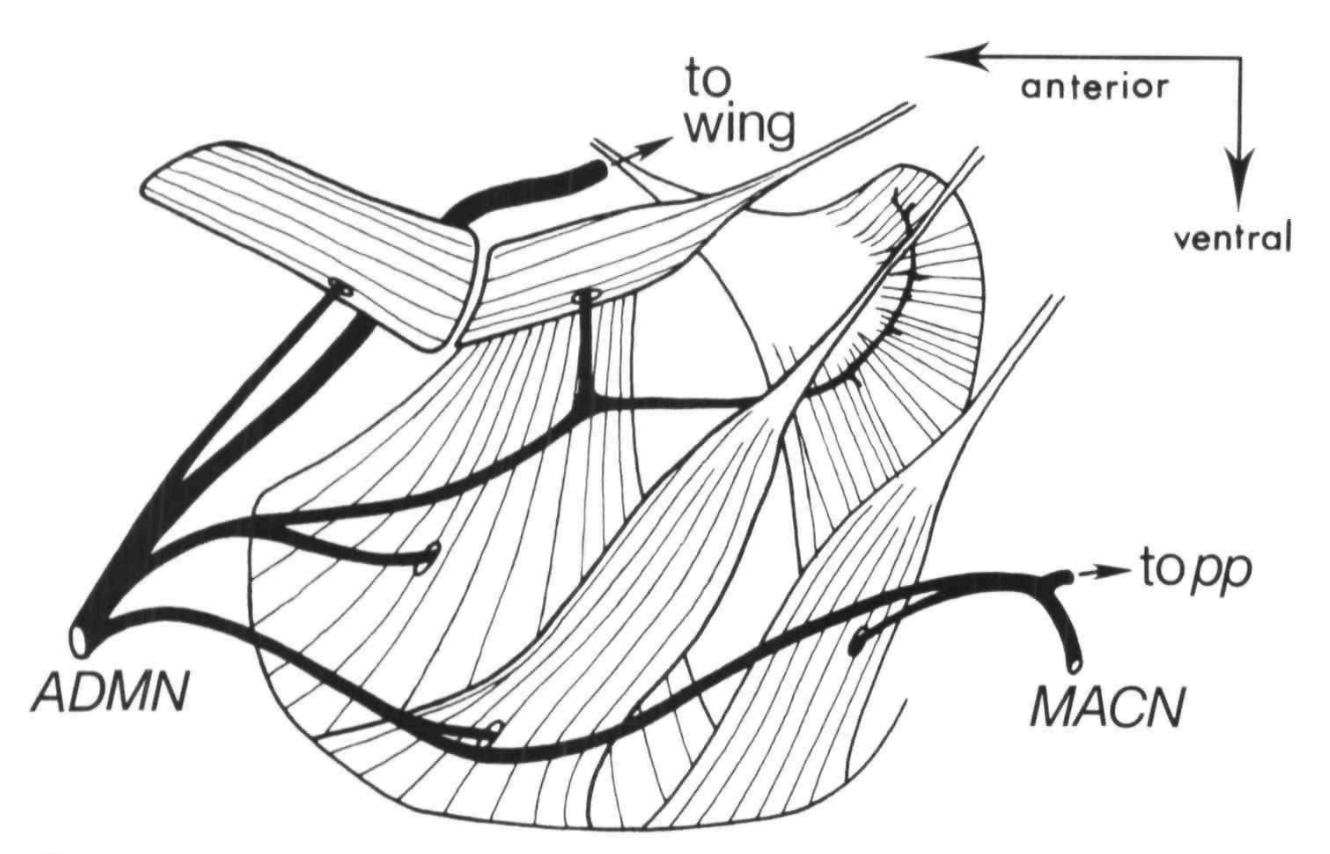
(1983d) King, D.G. and M.A. Tanouye Anatomy of motor axons to direct flight muscles in Drosophila. Journal of Experimental Biology 105: 231-239.
While working as a postdoc in Robert Wyman's lab at Yale, I had been delighted to discover that individually identifiable motor axons in a small animal the size of Drosophila could be reliably traced through serial thin sections all the way from their neuromuscular junctions into the thoracic central nervous system. This report of my histological observations was submitted jointly with Mark Tanouye, along with a companion paper (1983c, below) reporting Tanouye's recordings of electrical activity from the innervated muscles.
(1983c) Tanouye,
M.A. and D.G. King Giant fiber activation of direct flight muscles
in Drosophila. Journal of Experimental Biology 105: 241-251.
(1983b) King, D.G. and K.L. Valentino On neuronal homology: A
comparison of similar axons in Musca, Sarcophaga and Drosophila (Diptera: Schizophora).
Journal of Comparative Neurology 219: 1-9.
[Abstract]
Establishing homology at the level of individual cells is fraught with both technical and conceptual difficulties. This article
discusses these challenges while reporting putative homology for a set of axons individually identified
in three fly species from three different taxonomic families. This was accomplished primarily by demonstrating that their similarity
extended to the level of synaptic location. Unpublished continuation of this work has demonstrated the same configuration of synapsing axons
in several additional fly species. At the time this work was begun, Karen Valentino was an undergraduate student in
Robert Wyman's lab at Yale.
While working with Wyman as a postdoc, I offered her technical guidance as she prepared images of one of the specimens which
eventually appeared in this report.
(1983a) King, D.G. Evolutionary loss of a neural pathway from the
nervous system of a fly (Glossina morsitans, Diptera). Journal of Morphology
175: 27-32. [Abstract]
Two companion papers were submitted jointly with Mark Tanouye, a friend from our time together as postdocs in
Robert Wyman's lab at
Yale. This one, with Tanouye as principal author, reported physiological recording from Drosophila flight muscles.
My contribution to this paper was histological identification of sites where dye had been injected during recording. The other
paper of the pair (1983d, above) described the axons innervating those muscles.

Together with my paper "On neuronal homology..." (1983b, above), this publication represented
my initial foray into comparative study and evolutionary thinking.
My interest in an evolutionary perspective had been kindled by
Steven Jay Gould's monthly essays in Natural History magazine, under the heading
"This view of life."
(1982b) King, D.G., N. Kammlade, and J. Murphy A simple device to help reembed plastic sections. Stain Technology 57: 307-311. [Abstract]
Much of my histological study of flies involved relatively thick (2-5μm) serial sections of plastic-embedded specimens. With some help from the gadget described in this report, selected sections could then be lifted off their glass slides and re-sectioned for electron microscopy. (Ideally, as many as twenty sections for electron microscopy might be cut parallel to the face of one 2μm thick section.)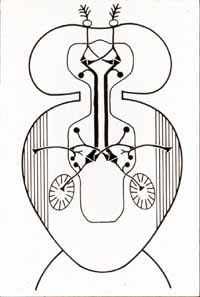
(1980) King, D.G. and R.J. Wyman Anatomy of the giant fiber pathway in Drosophila: I. Three thoracic components of the pathway. Journal of Neurocytology 9: 753-770. [Abstract]
This publication resulted from work completed during my post-doctoral experience (1976-1977) with Robert Wyman at Yale, where my attention was redirected from examining identified nerve cells in lobster to investigating the unique giant axons of Drosophila. The Roman numeral in the title betrays my optimistic but mistaken expectation that an additional publication would soon follow.My "prize" during this project was the discovery of the site of synapses between axons of the giant fiber pathway and motor axons to the fly's dorsal longitudinal muscle fibers. I was quite impressed that these synapses could be readily relocated in every fly specimen examined, thus demonstrating that the concept of identifiable individual nerve cells could even be applied at the level of synapses (see 1983b).
Wyman's lab was focussed on the Drosophila giant fiber pathway in hopes of identifying genes responsible for such conspicuously differentiated nerve cells. Imagining that phylogenic analysis might help illuminate the underlying genetics, I began looking for giant axons in other fly species, beginning with Musca and Sarcophaga. I subsequently collected images from 62 other species, a few of which were eventually published in a 2013a essay entitled "What can giant axons tell us about genetics and evolution?"
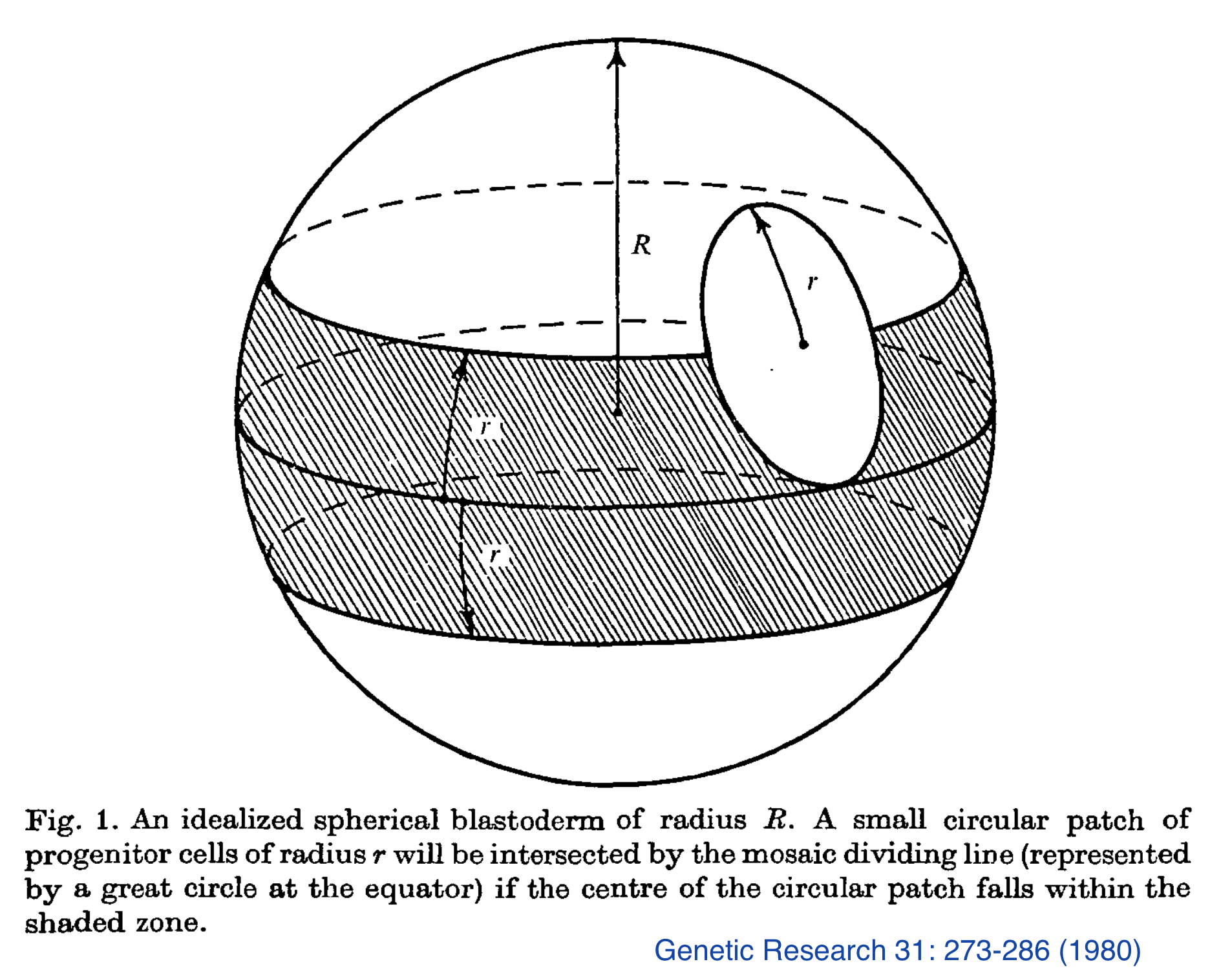
(1978) Wyman, R.J. and D.G. King, The dispersion of imaginal disc progenitor cells in the Drosophila blastoderm. Genetic Research 31: 273-286. [Abstract]
My postdoctoral mentor, Robert Wyman, was the principal author for this paper. My contribution -- a simple mathematical analysis of a simplistic model of the formation of genetic mosaics in Drosophila -- included a hand-inked diagram (at right) which inadvertently resembled the "death star" in the 1977 movie Star Wars.
(1976c) King, D.G. Organization of crustacean neuropil: II. Distribution of synaptic contacts on identified motor neurons in lobster stomatogastric ganglion. Journal of Neurocytology 5: 239-266. [Abstract] [email for PDF: dgking@siu.edu]
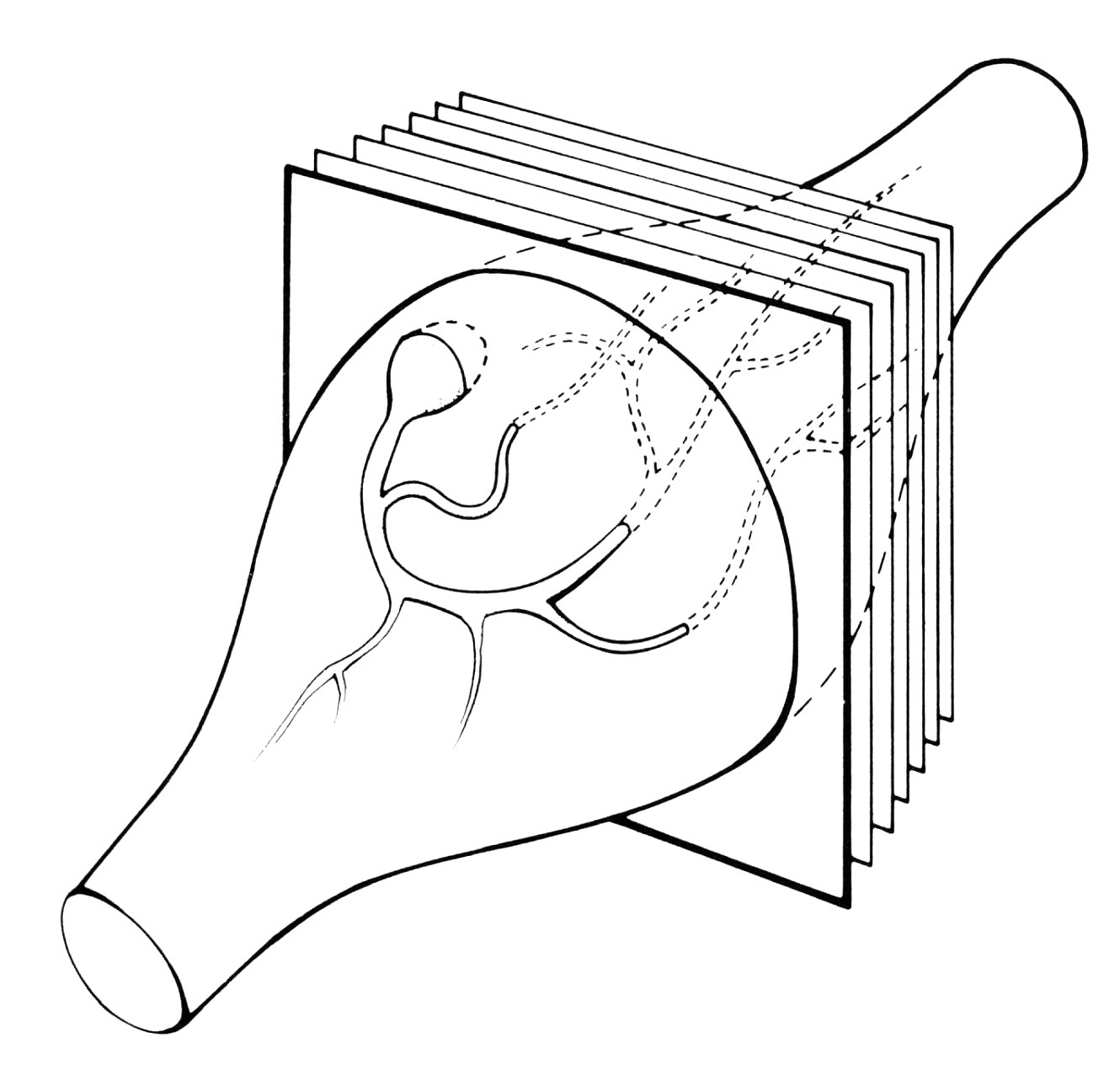
This paper, together with its companion paper (1976b, below), comprised my doctoral dissertation in neuroscience at UCSD. (I was tickled pink when this work was noticed in Scientific American [February 1978, Gordon M. Shepherd, "Microcircuits in the Nervous System"], a magazine I had read since childhood.)
This report is based on a single specimen whose cells had been individually identified electrophysiologically by my dissertation advisor, Allen Selverston. Serial-sectioning this specimen for electron microscopy enabled reconstruction of the shape of each cell's principal branches, including localization of specific synapses. Cutting, staining, and imaging approximately 5000 sections through this small piece of tissue (less than a millimeter in length), together with analyzing the resulting micrographs, was by far the most strenuous single piece of lab-work in my career.
☞ The phrase "identified motor neurons" in this paper's title refers to uniquely identifiable nerve cells. This concept -- still fairly new in the 1970s and remaining relatively unfamiliar even into the 21st century -- became a guiding theme in my subsequent research. Without realizing it at the time, I was both lucky and privileged to have been accepted as a student into an institution where "one of the major revolutions in neuroscience" was underway! I was first introduced to this astonishing concept when, as a brand-new graduate student in Ted Bullock's laboratory, I was assigned a project with a particular cell, the parabolic burster in the abdominal ganglion of Aplysia.
(1976b) King, D.G. Organization of crustacean neuropil: I. Patterns of synaptic connections in lobster stomatogastric ganglion. Journal of Neurocytology 5: 207-237. [Abstract] [email for full-text PDF: dgking@siu.edu]
Remarkably, this paper is still being cited 50 years after publication. Together with its companion paper (1976c, above), this paper comprised my doctoral dissertation in neuroscience at the University of California San Diego, completed with extensive support through the laboratory of Allen Selverston.
(1976a) Selverston, A., D.R. Russell, J.P. Miller, and D.G. King, The stomatogastric nervous system: Structure and function of a small neural network. Progress in Neurobiology 7: 215-290.
For several decades, a number of laboratories around the world have been intensively investigating the crustacean stomatogastric nervous system -- a model system for analyzing the mechanisms of motor pattern generation. I completed my own graduate study at one center for such study, Allen Selverston's lab at UC San Diego. This publication is a review of work up to 1975. [More recent reviews appeared in 1987, and 2016.] My principal contribution (in addition to assisting with overall editing) was a single-chapter preview of my doctoral dissertation, published separately and in greater detail in the two 1976 publications listed immediately above. (Coauthors Russell and Miller were also graduate students in Selverston's lab.)
until 1980 | 1981-1990 | 1991-2000 | 2001-2010 | 2011-2020 | since 2020 | Epilogue
 Comments and questions: dgking@siu.edu
Comments and questions: dgking@siu.edu
SIUC / Zoology / David King
https://dgkinglab.siu.edu/PUBLIST.HTM
Last updated: 28 April 2026 / dgk



{kind=link}