ZOOL 304, Class Notes
Hardy-Weinberg Frequencies
The Hardy-Weinberg Theorem is a mathematical idealization that serves
as a useful starting point for thinking about population genetics. Basically,
the theorem states that under certain idealize conditions, both allele frequencies
and genotype frequencies remain unchanged from one generation to the next.
A fundamental basis for this theorem is a mathematical relationship between
genotype frequencies and allele frequencies.
We consider the simple case of a diploid population with one locus.
At this locus we consider only two alleles, A and a.
[The basic concepts are the same with more than two alleles,
just a bit more complicated. Besides, no matter how many alleles
occur, one may always pick one allele, say A, and then lump together
all other alleles as "not A".]
We label the allele frequencies as p and q, thus:
frequency of A = p We abbreviate this
as [freq A]
frequency of a = q We abbreviate this
as [freq a]
And the genotype frequencies (sometimes called the Hardy-Weinberg frequencies)
are:
[freq AA] = p2
[freq Aa] = 2pq
[freq aa] = q2
Since this relationship is a source of some difficulty for many students,
this page offers a review explanation, continuing below.
For further explanation of the Hardy-Weinberg Theorem itself, see Mathematical
Models.
First, a bit about those "p"s and "q"s.
By convention, the letter "p" is commonly used to represent
the frequency of one allele (usually labelled "A") in a population,
while "q" stands for frequency of the alternative allele
(labelled "a").
A frequency, of course, is just a proportion (or fraction) of the
total.
We use the word "frequency", rather than "proportion",
when measuring countable items like alleles in a gene pool. But our
math is done just as as if a frequency were a continuous variable like a
proportion.
Thus, for the particular gene under consideration in our simple two-allele
population, p is just the fraction of the genes in the population which
are of allele type A while q is the fraction with allele type
a. Since our simple example treats a population with only the
two alleles A and a, the sum of these two fractions equals one,
the total of all the alleles.
p + q = 1
Example: Suppose you order a pizza topped with pepperoni
on one quarter and quince on the remaining three quarters.
(Quince are a kind of fruit; yes, it's a silly example, but it keeps the
ps and qs in order.) Then you cut the pizza into eight
slices. The frequency of slices with pepperoni is 2
out of 8 or 1/4 while the frequency of slices with quince
is 6 out of 8 or 3/4. The sum of pepperoni plus quince
is 1/4 + 3/4 = 1 or 2 slices plus 6 slices equals 8 slices, which equals
on entire pizza. p + q = 1.
The probability of choosing an allele at random is closely related
to the frequency of that allele. Basically, for a sufficiently large
randomly mating population, probability equals frequency.
Hopefully this is fairly obvious. If not, just imagine
that alleles in the gene pool are marbles in a bag. If the bag contains
75 red marbles and 25 blue marbles, the frequency of red marbles is just
75 out of 100 (or 0.75, or 3/4) and the probability of reaching into the
bag and picking a red marble is also 75 out of 100 (or 0.75, or 3/4).
If you don't like thinking of alleles as marbles, think
of them more biologically as types of gametes. Under the assumptions
of the Hardy-Weinberg model, mating is a process equivalent to stirring
into one big pot all of the gametes (without distinguishing eggs and sperm)
produced by the population. Any particular zygote forms as the random
combination of two gametes, equivalent to reaching into the pot and choosing
each one at random.
(What about more than two alleles? Everything said here may be modified
to accommodate more than two alleles, just by introducing additional variables
for the frequencies of the additional alleles, such that the sum of all the
frequencies must always be 1. Alternatively, even if there are
several alleles, one may always pick one allele, say A, and then lump
together all the rest as as "not A".)
Explanation of genotype frequencies based on elementary algebra.
The mathematic basis for the genotype frequencies, AA : Aa : aa =
p2 : 2pq : q2,
is algebraic and is based on the simple identity,
p + q = 1
(This fact itself should be easy to remember -- just recall that the parts
must equal the whole. That is, the frequencies of the elements which
comprise a population must sum to one.)
It is also sometimes helpful to remember that p = 1 - q and
q = 1 - p . These are derived by simple algebra
from p + q = 1.
Each genotype represents a random pairing of two alleles. Since frequencies
(and probabilities) of independent events taken together combine by multiplication,
we get the genotype frequencies by simple polynomial multiplication. Thus:
( p + q ) x ( p + q ) = p2
+ 2pq + q2
So, there they are. The genotype frequencies are simply the terms
in the algebraic expansion of ( p + q )2 .
If algebra is not your strength, this identity can also be interpreted geometrically,
as follows.
Explanation of genotype frequencies based on elementary geometry.
Do you recall from grade school that you find the area of a rectangle by
multiplying the lengths of the two sides?
Believe it or not, that's all you need to know to figure out genotype frequencies
from allele frequencies. .
First, let's begin by seeing how multiplying the sides of a rectangle
to find its area is just like polynomial multiplication in algebra.
|
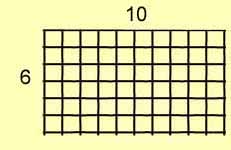
For example, consider a rectangle with sides of 6 and 10. The area,
of course, is:
6 x 10
= 60
|
 |
|
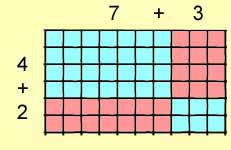
Now consider that each side may be subdivided. For example, 6 =
( 4 + 2 ) and 10 = (7 + 3 ).
6 x 10 = ( 4 + 2 ) x
( 7 + 3 )
= (
4 x 7 ) + ( 4
x 3 ) + ( 2 x 7
) + ( 2 x 3 )
= 28
+ 12 + 14 + 6
= 60
|
 |
|
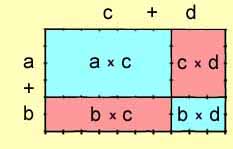
More generally, any rectangle may be composed of four smaller rectangles,
each the product of segments along its sides.
( a + b ) x
( c + d ) =
( a
x c ) + ( c x
d ) + ( b x
c ) + ( b
x d )
|
 |
Now let's apply this geometric multiplication to population genetics. Just
imagine all the individuals comprising a population to be arranged into a
square array. Our tool for doing this is the Punnett
Square.
(Here is an irrelevant mathematical
aside on the Pythagorean theorem, just for fun.)
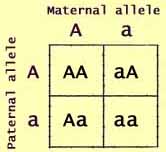
The Punnett Square
The Punnett square was introduced by English geneticist Reginald
Crundall Punnett as a tool for visualizing genetic combinations.
In its simplest form, a Punnett square shows how alleles combine to form
genotypes.
|
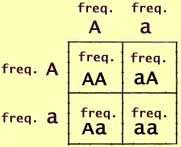
Along one side of the square are listed the alleles that a zygote may
receive from its mother. Along the other side are listed the alleles
that the zygote may receive from its father. Within each square
are listed the resulting genotypes.
|
 |
|
However, instead of just listing alleles and genotypes, the sides of
our square can represent the allele frequencies. Then the boxes
represent the genotype frequencies.
But now this becomes more than a simple listing. The square can
now be used for computation.
|
 |
|
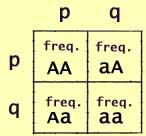
We can now substitute algebraic or numerical representations of allele
and genotype frequencies.:
p is the frequency of A and
q is the frequency of a.
|
 |
|
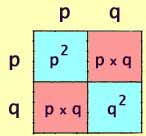
Finally we use multiplication by geometry, just
like in the rectangle above, to see that:
[freq AA] = p2
[freq Aa] = 2pq
[freq aa] = q2
|
 |
|
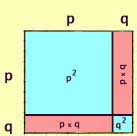
This may appear just a bit more intuitive if we make the segments along
the sides of the square proportional to the allele frequency.
As shown at right, p = 0.8 and q = 0.2
)
|
 |
Notes for chapter 1 / 2
/ 3 / 4 / 5
/ 6 / 7 / 8
/ 9 / 10 / 11
/ 12 / 13 / 14
/ 15 / 16 / 17
304 index page
The utility of algebra-by-geometry is not limited to Punnett Squares. Here
is an elegant proof of the Pythagorean Theorem from Euclidean geometry.
You remember, "In any right triangle, the square of the hypotenuse
is equal to the sum of the squares of the other two sides."
|
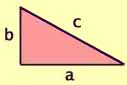
Take any right triangle with sides a and b
and hypotenuse c.
Area = 1/2 x ab
|
 |
|
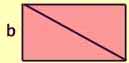
Put two of these triangles together to form a rectangle.
Area = 2 x (1/2 x ab)
|
 |
|
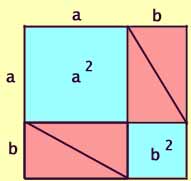
Next, arrange two such rectangles into a larger
square with side a + b.
Clearly, the area of this larger square is equal to four of the triangles
plus the two blue squares.
Area = (a + b)2
= a2 + b2 + [4 x (1/2 x ab)]
|
 |
|
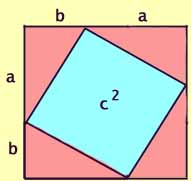
Now rearrange the four pink triangles within the same larger
square. The side of the outer square is still (a + b),
so the area of the outer square also still (a + b)2.
Area =
= c2 + [4 x (1/2 x ab)]
Since the larger square has not changed its size (side a + b),
and there are still four pink triangles, the blue areas must also remain
equal. Thus the sum of the areas of the two blue squares in the
diagram above must equal area of the single blue square in this diagram.
Therefore,
a2 + b2 = c2
|
 |
In words, the square of the hypotenuse ( c2 ) is
equal to the sum of the squares
(a2 + b2 ) of the other two sides.
Now, back to evolution...
Notes for chapter 1 / 2
/ 3 / 4 / 5
/ 6 / 7 / 8
/ 9 / 10 / 11
/ 12 / 13 / 14
/ 15 / 16 / 17
304 index page
Comments and questions: dgking@siu.edu
Department of Zoology e-mail: zoology@zoology.siu.edu
Comments and questions related to web server: webmaster@science.siu.edu
SIUC / College
of Science / Zoology / Faculty
/ David King /
ZOOL 304
URL: http://www.science.siu.edu/zoology/king/304/h-w.htm
Last updated: 13 October 2003 / dgk